


Introduction
The autophagy maintains the homeostasis by binding the intracellular organelles and protein to lysosome and thereby degrading them when the nutrient supply to the cells is blocked. The major functions of autophagy include self-control, response to the stress and the alternative measures for energy and survival. But if this is persistently present, this would lead to a complete disruption of the cellular morphology. These phenomena are referred to as type II programmed cell death or intracytoplasmic cell death. It is known that the cancer cells supply the nutrients through the autophagy but this leads to the apoptosis when exposed to the long-standing stimuli [1,2].
One of the anti-cancer drugs that are commonly used to treat patients with gynecological cancer, paclitaxel has an anti-cancer effect are based on the apoptosis [3]. But most of the patients with progressive or recurrent cancer have a lower rate of treatment response to these anti-cancer drugs. To overcome this, efforts have been made to raise the treatment response by administering concomitant anti-cancer drugs rather than single one and introducing new anti-cancer drugs. To date, these efforts have revealed the limitations to the applicability to a clinical setting.
On the other hand, it is known that the phosphatidylinositol 3 phosphate kinase (PI3K)/Akt/mammalian target of rapamycin (mTOR) signaling pathway as well as the apoptosis is involved in the mechanisms by which the autophagy occurs. As described here, if there are any chances that these two involved mechanisms for apoptosis might be regulated, this would further lead to the programmed cell death. In this study, we examined whether there is a possibility to maximize the treatment effect using rapamycin, an inducer of the autophagy, concomitantly with conventional types of apoptosis-based anti-cancer drugs in a model of uterine cervical cancer.
Materials and Methods
1. Culture of HeLa cell lines and the treatment of them with drugs
The HeLa uterine cervical cancer cell lines were cultured in a Dulbecco's modified Eagle's medium containing 10% fetal bovine serum (FBS) in conditions of 37Ōäā and 5% CO2. 1├Ś104/mL HeLa cell lines were cultured on a 96-well plate (Nunc, Rochester, NY, USA) for 48 hours. This was followed by the treatment with an anti-cancer drug, paclitaxel (Sigma-Aldrich, St. Louis, MO, USA), and rapamycin (Sigma-Aldrich), an inducer of autophagy.
Following the culture of HeLa uterine cervical cancer cell lines, they were treated with paclitaxel at varying concentrations of 0, 5, 10, 15, and 20 nM for 24 hours. This was followed by the examination of their inhibitory effects on cell survival. The cell survival was examined 24 hours after the treatment with rapamycin at varying concentrations of 0, 5, 10, 15, and 20 nM.
On the other hand, to examine the synergistic effects of two drugs in increasing the survival of cells, we administered paclitaxel at varying concentrations of 0, 5, 10, and 20 nM to the HeLa cells that had been pre-treated with rapamycin 10 nM for three hours. Then, the cell viability was examined.
The assessment was done with the methods for measuring 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide (MTT, Colorimetric assay kit, Chemicon Inc., Temecula, CA, USA) which had been modified from tetrazolium based colorimetric assay. Following the addition of MTT samples 20 mg/mL, the culture was done for four hours. After the removal of culture medium, followed by the addition of dimenthylsulfoxide (DMSO), an analysis was done at a wave-length of 450 nm.
2. The quantitative analysis of the apoptosis and autophagy due to paclitaxel in HeLa uterine cervical cancer cell lines
1-5├Ś105 HeLa uterine cervical cancer cells were cultured for a day and then treated with 5 nM paclitaxel. Twenty-four hours after the treatment with paclitaxel, both paclitaxel-treated cells and their non-treated controls were administered trypsin. Then, the cells were placed in phosphate buffered saline (PBS) containing 1% paraformaldehyde. This was followed by the fixation with 70% cool ethanol. Annexin V propidium iodide (PI) staining were done with ApoDETECT Annexin V-FITC kit (Invitrogen, Calsbad, CA, USA). According to the manufacturer's instructions, changes in the apoptosis were examined. The stained cells were subjected to a quantitative apoptosis assay using a flow cytometry (FACS Vantage SE, BD Bioscience, San Jose, CA, USA). Thus, the cells undergoing apoptosis were counted.
For the quantitative analysis of autophagy, changes in the development of acidic vesicular organelles were examined. That is, 5├Ś105 HeLa cells were cultured overnight. This was followed by a 24-hour treatment with 5 nM paclitaxel. Paclitaxel-treated cells and their non-treated controls were stained with acridine orange (Sigma-Aldrich) at a concentration of 0.5 ug/mL at 37Ōäā for 15 minutes with the sunlight blocked. The cells were rinsed with PBS twice and then treated with trypsin. Then, the cells were suspended on PBS containing 1% FBS, whose number was counted using a flow cytometry. This was followed by the analysis using CellQuest 7.0 software (Beckman Coulter Co., Brea, CA, USA).
3. An assay of the proteins undergoing apoptosis and autophagy
1├Ś106/mL cells were cultured on a 100 mm2 dish for 48 hours and then harvested. This was followed by a 30-minute centrifugation at a temperature of 4Ōäā and at 12,000 rpm. The extracted cells were reacted in a 100 ┬ĄL lysis buffer at 4Ōäā for 30 minutes and then dissolved. The cell lysate was centrifuged at a temperature of 4Ōäā and at 12,000 rpm. Thus, the protein was harvested. This was followed by the measurement of the concentration of protein using Bio-Rad Protein Assay (Bio-Rad, Philadelphia, PA, USA).
The protein 50 ┬Ąg was boiled for 5 minutes and then subjected to a 2-hour electrophoresis using a 10% sodium dodecyl sulfate/polyacrylamide gel at 100 V. Then, the protein was transferred to a nitrocellulose paper at 360 mA for an hour. This was followed by the blockage of unnecessary factors from 5% skim milk and 0.05% tris buffered saline with Tween 20 (TBS-T) solution on the membrane for an hour.
Using the primary antibodies against caspase 3 (Santa Cruz Biotechnology, Santa Cruz, CA, USA) and light chain (LC) 3 (Santa Cruz Biotechnology), the membrane was reacted overnight at 4Ōäā. The membrane was rinsed with the tris-buffered saline and TBS-T, three times. Each membrane was reacted with the secondary antibody (anti-mouse IgG, Santa Cruz Biotechnology) at room temperature for an hour. Then, the membrane was rinsed with TBS-T three times. The specific protein bands were obtained using the ECL Western blotting system (Amersham, Piscataway, NJ, USA).
4. LC3 transfection
For the quantitative analysis of autophagy, the cells were labeled with green fluorescent protein (GFP)-LC3. Then, changes in the number of spots were examined using a confocal microscope (quantitative autophagic assay).
HeLa cells were transiently transfected into pCMV-GFP-LC3 expression vector (Cell Biolab, San Diego, CA, USA). Then, following the treatment with 10 nM rapamycin and 5 nM paclitaxel for 24 hours, the cells were fixed with 4% formaldehyde. The cells were rinsed with PBS and then examined using a polarized microscope (Olympus FluoView 1000, Olympus, Melville, NY, USA). At a magnification of ├Ś200, we counted and then compared the number of cells where more than ten GFP-LC3 spots were stained.
Results
1. Anti-cancer effects of paclitaxel (an anti-cancer drug) and rapamycin (a regulator of the autophagy) on the HeLa cells
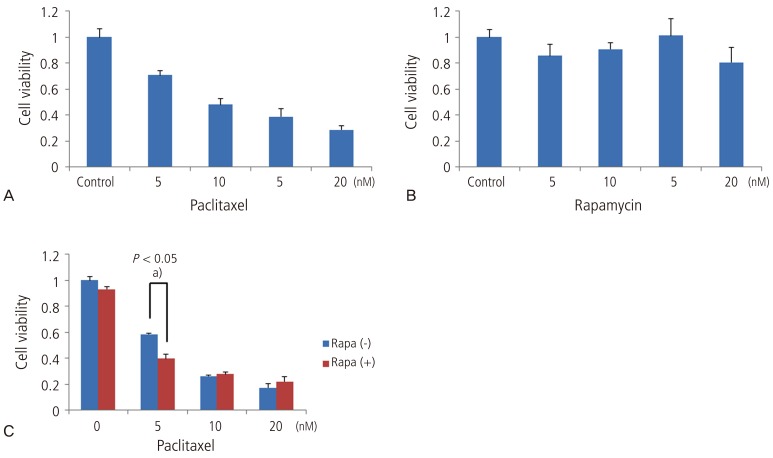
HeLa uterine cervical cancer cell lines were treated with paclitaxel at varying concentrations. Then, we determined the 50% inhibition concentration (IC50) where the cytotoxicity reached the maximum level at 24 hours. The cell survival was inhibited in a concentration-dependent manner following a 24-hour treatment with paclitaxel at varying concentrations of 5, 10, 15, and 20 nM. In addition, the IC50 of paclitaxel, at which the growth of HeLa cells is decreased by 50%, ranged between 5 nM and 10 nM (Fig. 1A).
The HeLa cells were treated with rapamycin (a regulator of the autophagy) at varying concentrations of 5, 10, 15, and 20 nM for 24 hours. But there were no concentrations of rapamycin at which the cytotoxicity appeared (Fig. 1B).
In addition, we also examined the effects of concomitant treatment with paclitaxel and rapamycin at varying concentrations in inhibiting the cell survival. Following a 3-hour pre-treatment with 10 nM rapamycin, the cells were then treated with paclitaxel at concentrations of 0, 5, 10, and 20 nM. Then, 24 hours later, it was examined whether there were inhibitory effects on the survival of HeLa uterine cervical cancer cells. This showed that montherapy with rapamycin had no effects on the cell survival. In addition, following the pre-treatment with 10 nM rapamycin, the cells were treated with paclitaxel at concentrations of 10 nM and 20 nM. This showed that there were no significant differences in the inhibitory effects on the cell survival as compared with a lack of the pre-treatment with rapamycin. With the pre-treatment with rapamycin followed by the treatment with 5 nM paclitaxel, the degree of inhibitory effects on the cells survival was higher as compared with a lack of the pre-treatment with rapamycin. This difference reached a statistical significance (P<0.05) (Fig. 1C).
The difference in the cytotoxicity, described herein, can be explained by the synergistic effects of rapamycin (an inducer of the autophagy) at a certain concentration of paclitaxel.
2. The quantitative analysis of apoptosis and autophagy
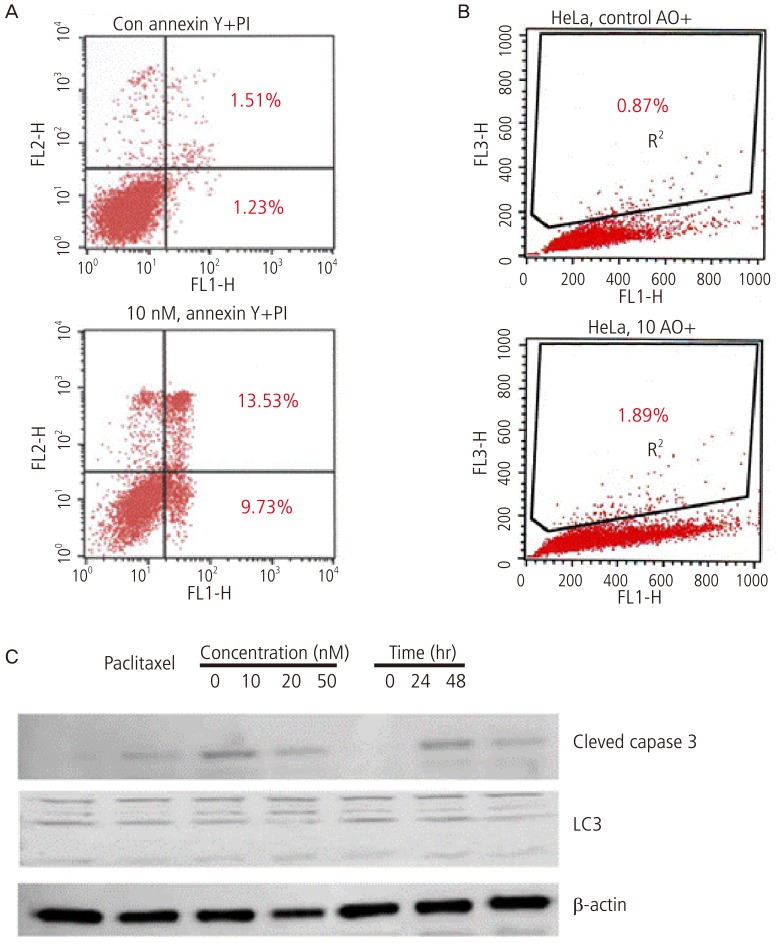
It was examined whether paclitaxel-induced inhibitory effects on the cell survival originate from the apoptosis and autophagy. To examine the mechanisms by which the cell survival is inhibited at 24 hours following the treatment of HeLa cells with 10 nM paclitaxel, the cells were stained with annexin V. The staining property was examined using a flow cytometry (BD FACS). Thus, it was confirmed that the apoptosis was involved in the mechanisms (Fig. 2A). It was also shown that the proportion of cells undergoing apoptosis was increased from 2.74% to 23.26% as compared with prior to the treatment with paclitaxel.
Moreover, to examine the quantitative changes in the autophagy, the cells were treated with 10 nM paclitaxel and then stained with acridine orange within 24 hours thereafter. Then, the number of acridine orange-positive cells was counted using a flow cytometry. This showed that the proportion of cells undergoing the autophagy was increased from 0.87% to 1.89% as compared with prior to the treatment with paclitaxel. It was also shown that the paclitaxel monotherapy had no effects on the autophagy (Fig. 2B).
To identify the proteins that are involved in the apoptosis and autophagy, HeLa cells were treated with paclitaxel at varying concentrations of 0, 10, 20, and 50 nM for 24 hours. Following the treatment with 10 nM paclitaxel for 24 hours and 48 hours, the degree of the expressions of caspase 3 and LC3 was measured using a western blot analysis. Cleaved caspase 3, involved in the apoptosis, showed an increased degree of expression as the concentration of paclitaxel was elevated. But there were no changes in LC3 protein that is involved in the autophagy (Fig. 2C).
3. LC3 transfection
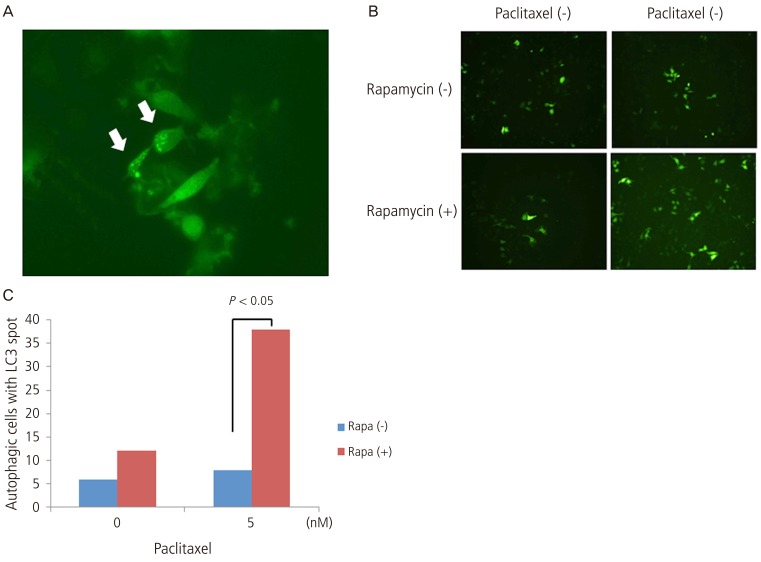
After the LC3 transfection into the HeLa cells, followed by the treatment with paclitaxel and rapamycin, changes in the cells were examined using a light microscopy. Once the autophagy occurs, LC3 binds to the autophagosome. This leads to the formation of spots, which is a key marker of the autophagy (Fig. 3A). There was no increase in the number of cells where more than ten LC3 spots appeared following the treatment with 5 nM paclitaxel. Following the treatment with 10 nM rapamycin, however, it was increased. With the concomitant administration of two drugs, it was further increased (P<0.05) (Fig. 3B, C).
Discussion
In cases of nutrient deficiency, the tumor cells are dependent on the autophagy for their survival in the early stage of tumorigenesis. If this is continued, it would lead to the apoptosis. But this remains obscure at the present. In addition, it also remains unclear whether the autophagy directly causes the apoptosis or it is simply a phenomenon that occurs secondarily to the apoptosis [1,2].
As the proteins involved in the autophagy, beclin 1 was first identified as a substance that binds to Bcl-2 (oncogene) based on the yeast two hybrid methods. It plays a role in triggering the occurrence of autophagy. Earlier studies have reported that the expression of beclin 1 led to the occurrence of autophagy and this eventually inhibited the tumor occurrence [4,5]. It has been shown that the inhibition of beclin 1 expression promoted the development of several types of tumor. Thus, it was confirmed that beclin 1 is a tumor suppressor gene. Moreover, it has also been shown that the autophagy is involved in the suppression of tumor occurrence [6,7]. Recent studies have also shown that some tumor suppressor genes such as the phosphatase and tensin homologue (PTEN) and TSC1/2, as well as beclin 1, are also involved in triggering the occurrence of autophagy [8,9]. The proposed mechanisms by which the autophagy is involved in the suppression of tumor occurrence are as follows: 1) The degradation of the key protein and cell organelles that are essential for suppressing the cell growth. 2) The production of reactive oxygen species and the removal of damaged cell organelles causing the genotoxic stress. 3) The induction of the occurrence of autophagy, another programmed cell death than the apoptosis, leading to the occurrence of apoptosis [10].
It is widely known that the PI3K/Akt/mTOR signaling pathway is involved in the molecular biological mechanisms by which the autophagy occurs [8]. In the presence of the stimulation of growth factors, the growth factor receptors activate type I PI3K on the plasma membrane and thereby inhibits the transformation of autophagy. PI3K stimulates the Akt, the downstream target, and activates mTOR. Thus, it suppresses the p70S6 kinase-induced autophagy. On the other hand, if the PTEN gene is over-expressed, this would cause an autophagy with the antagonistic effects of type I PI3K. Ras two effects. If it stimulates the PI3K, this leads to the suppression of autophagy. If it selectively stimulates the RAF1-mitogen activated protein kinase (MEK)-extracellular signal-regulated kinase (ERK), this triggers the occurrence of autophagy. The complex between rapamycin (mTOR inhibitor), type III PI3K and beclin 1 stimulates the autophagy. The differential features of autophagy-induced type II programmed cell death from type I one are not only that the apoptosis is morphologically characterized by changes in chromatin density, the formation of DNA laddering or fragmentation but also that the autophagy is characterized by an abundant presence of the intracellular autophagic vacuoles, the degradation of Golgi apparatus, ribosome and endoplasmic reticulum. In addition, it has also been known that the autophagy leads to a programmed cell death that occurs in a non caspase-dependent manner. But both events are involved in the PI3K/Atk/mTOR signaling pathway that is essential for the tumorigenesis. It is therefore presumed that the molecular biological mechanisms of programmed cell death might be inter-related.
There are some reports confirming that z-VAD (caspase inhibitor) triggers the occurrence of autophagy rather than apoptosis [11]. In addition, it has also been reported that the apoptosis is inhibited following the treatment with rapamycin (an inducer of the autophagy) [12]. Still, however, it remains unclear whether the autophagy would be helpful for the apoptosis during the anti-cancer treatment or it would have an antagonistic effect.
In Korea, the uterine cervical cancer is one of the most common gynecological cancers, most of which are treated with surgery and radiotherapy. It is known as a carcinoma which is sensitive to platinum anti-cancer drugs such as cisplatin or taxanes such as paclitaxel [13]. Recently, there is a decreasing tendency in the incidence of uterine cervical cancer with the help of the Korean Uterine Cervical Cancer Screening Program. It is also known that the treatment outcomes are so excellent that the rate of complete cure is relatively higher with the early detection of it.
Paclitaxel is one of the representative taxane anti-cancer drugs. It is known to stabilize the microtubules during the DNA synthesis and thereby to suppress the mitosis of the cancer cells. It has been tested on several clinical trials during the 1980's. In the 1990's, its indications for various types of cancer have been approved by the Food and Drug Administration. Recent studies have reported its excellent efficacy on not only uterine cervical cancer but also breast cancer, ovarian cancer, lung cancer, head-and-neck cancer and Kaposi's sarcoma. In cases of progressive cancer or recurrent uterine cervical cancer, however, it shows a lower degree of treatment effect. A poor prognosis is therefore unavoidable in these cancers. For this reason, the compound regimens are used to raise the efficacy of anti-cancer drugs. But side effects prevalently occur without markedly increasing the survival. This indicates that other treatment regimens than the anti-cancer drugs are necessary [14].
Recent studies have disclosed the mode of action of biological agents. Meanwhile, novel target treatment agents based on the inhibitors of monoclonal antibodies such as vascular endothelial growth factor and epidermal growth factor receptor have been actively introduced for the treatment of other cancers than uterine cervical cancer [15]. An inducer of the autophagy, rapamycin is also an inhibitor of mTOR; its efficacy on renal cancers has been demonstrated on several clinical studies conducted in cancer patients [16]. But rapamycin itself has an inhibitory effect on the cell growth to a greater extent than its cytotoxic effect. Therefore, its efficacy should be controlled with concomitant regimens with other drugs with a different mode of action rather than the monotherapy. In addition, the autophagy may induce the secondary apoptosis, but this is somewhat reversible and time-consuming. Presumably, it would therefore be recommended to use the adjuvant use rather than the direct anti-cancer effect of rapamycin [17].
Our results showed that rapamycin (an inducer of the autophagy) was effective in significantly increasing the autophagy in HeLa uterine cervical cancer cells. It was also shown that its degree of inhibitory effect was relatively higher as compared with the paclitaxel monotherapy. Besides, we also transfected LC3, an intermediate substance from autophagy using green fluorescent protein and then examined the quantitative changes. This showed that LC3 spots were increased at a lower concentration of paclitaxel. In addition, they were further increased when the autophagy was induced prior to the paclitaxel treatment. Therefore, our results showed that the use of an inducer of the autophagy could enhance the primary treatment response of conventional types of anti-cancer drugs in a model of gynecological cancer. In addition, it was also shown that the use of an inducer of the autophagy promoted the programmed cell death and this led to the increased anti-cancer effect.
At a certain concentration of paclitaxel, its concomitant use with rapamycin (a regulator of the autophagy) further increased an inhibitory effect on the cell survival (Fig. 1C). With the administration of 10 nM and 20 nM paclitaxel, there was a progression of the apoptosis. Therefore, we failed to confirm whether there are additional effects in inhibiting the cell survival when the survival of cells were suppressed at a proportion of 70% to 80%. With the administration of 5 nM paclitaxel at which the survival of cells was inhibited at a proportion of 50%, however, rapamycin additonally had an inhibitory effect on the cell survival at a proportion of approximately 15%. These results suggest that the autophagy would be helpful for the anti-cancer effects when the anti-cancer drugs such as paclitaxel are used in an in vivo setting if there are any chances that they could not sufficiently reach the target sites because the size of tumor is relatively larger or the blood vessels are not well developed.
Sun et al. [18] reported that the degree of cytotoxic effect of anti-cancer drugs was increased with the over-expression of beclin 1 in CaSki uterine cervical cancer cells. According to these authors, the toxicity of anti-cancer drugs such as cisplatin and paclitaxel increased and then amplified the autophagy of beclin 1 as well as apoptosis. Then, they noted that the autophagy play a key role in regulating the anti-cancer effects. According to Zou et al. [19], ARHI (a tumor suppressor gene) induced the autophagy and this led to a further increase in the inhibitory effects of paclitaxel in breast cancer cells. Recent reports have shown that the inhibitory effect of paclitaxel was increased following the use of siRNA transcripts of 3-methyladenine or beclin 1, both of which are inhibitors of the autophagy [20]. This implies that a regulator of the autophagy would be used as an adjuvant treatment strategy to enhance the efficacy of anti-cancer drugs.
In the current study, we attempted to raise the degree of treatment response in patients with uterine cervical cancer and to identify novel treatment modalities that can overcome the limitations of conventional types of anti-cancer drugs. To put this in another way, it has been hypothesized that the future treatment modalities will be effective in enhancing the early efficacy of the pharmacological treatments, preventing the recurrence of cancer and treating it by concomitantly using the target treatment agents or immune boosters based on other biological agents with a different mode of action from conventional types of standard anti-cancer therapies.
In conclusion, our results indicate that an inducer of the autophagy was effective in increasing the efficacy of conventional types of anti-cancer drugs causing an apoptosis in uterine cancer cells. Thus, our results provided an evidence demonstrating that an inducer of the autophagy might be used an adjuvant modality during the anti-cancer pharmacological treatments.






")











